最近の研究を少し整理して紹介します。
1. 競争関係にある多種の共存メカニズム
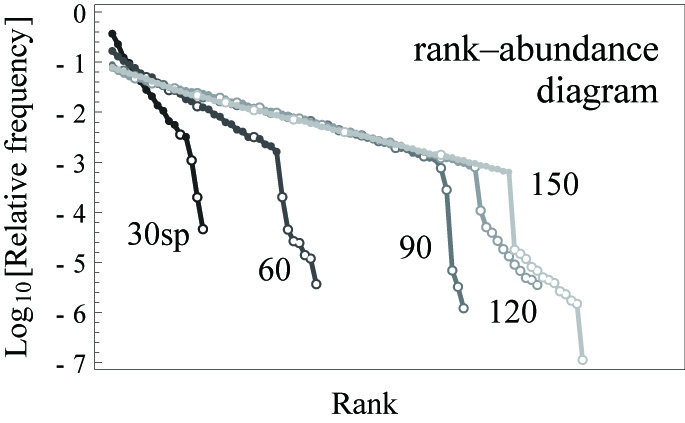
種間競争は生物群集に普遍的に存在する相互作用ですが、それは多種共存を抑制する要因として捉えられがちです。その種間競争に注目したモデルの一つに移住モデルがあります。このモデルは、生物の幼生や種子がコロニーを形成するためのサイトをめぐって競争する過程を定式化したものです。各種には、サイトをめぐる競争での優劣にもとづく順位関係があり、順位の高い種は自分より順位の低い種との競争に打ち勝ち、サイトを占めてコロニーを作れます。その一方で、競争力の低い種は幼生の生産力が高くなるというトレードオフがあるとします。このモデルでは多種が特徴的なランク-アバンダンス関係 (RAD) を示しながら共存することが可能です。その RAD の性質が、実際の生物群集でみられるランク-アバンダンス関係の特徴をよく説明することを示しました。このことは、実際の生物群集における多種共存が、種間競争によってもたらされている可能性を示唆します。
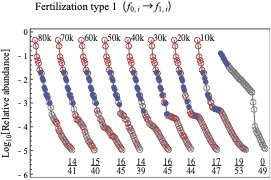
長期にわたる栄養添加が植物群集の構造を大きく変えることが、40年以上前に Kempton によってはじめて報告されました。その後、同様な現象が多くの研究者によって多様な群集について報告されています。それらの研究はいずれも、生物種数の減少、ランク-アバンダンス関係の傾きの増大、アバンダンスが大きい種の交代などの一貫した傾向を示しています。栄養添加に伴うこうした群集構造の変化は、植物の形質の変化に注目したり、ニッチ次元の変化と関連づけて議論されていますが、それらの反応が上記のような一貫した変化傾向をもたらすメカニズムは分かっていません。この問題に移住モデルを適用してメカニズムを論じました。競争力の低い種は幼生の生産力が高くなるというトレードオフがある状況で、栄養の添加が生産力に関する制約を緩めるとすると、上記のような一貫した群集構造の変化が起きうることを示しました。また、トレードオフの緩和のパターンによって、群集のバイオマスが変化する場合も変化しない場合もあり、このことも栄養添加がバイオマスに与える影響とよく一致します。
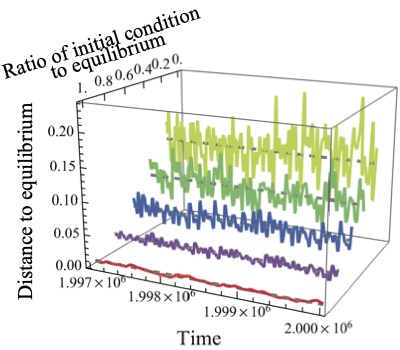
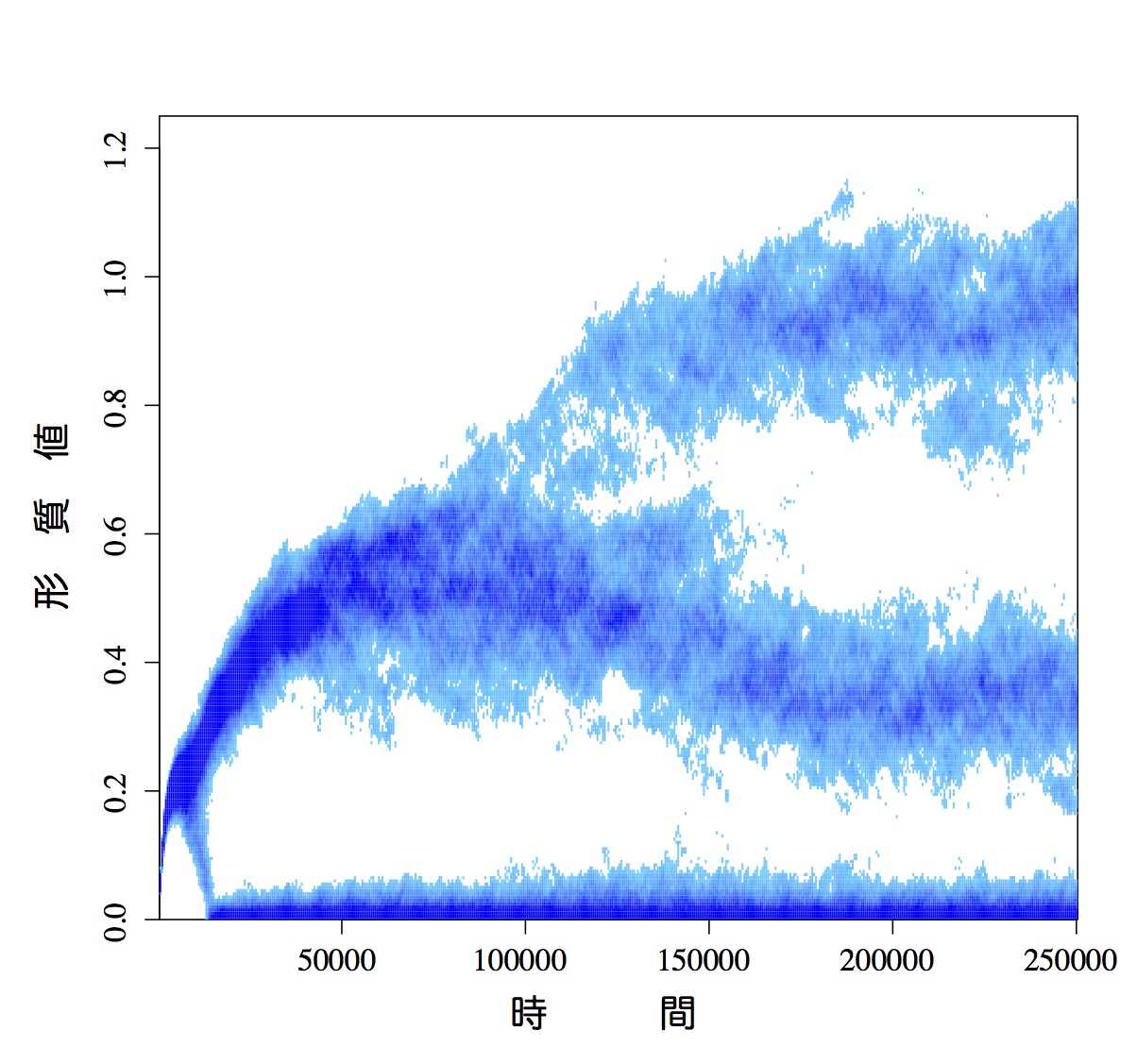
移住モデルでは、ある生物種の幼生とコロニーを形成するためのサイトとの出会いの確率を、幼生プール中におけるその種の幼生の密度の関数として記述します。この仮定を変更し、幼生とサイトとの出会いの確率を、幼生プール中におけるその種の幼生の密度および頻度の関数として、多種系ののダイナミクスを調べました。数値解析とシミュレーションを組み合わせた解析の結果、出会いの確率が完全に密度依存、あるいは密度と頻度の両方に依存する場合には安定な平衡状態が存在して、多種が安定的に共存する状態に収束します。それに対して完全に頻度依存の場合、ダイナミクスは永続的に不規則な変動を続け、しかもそうした軌道は初期状態に依存して無数に存在することが分かりました。それらの軌道の特性を調べてみると、不規則ではあるもののカオスではなく、strange non-chaotic attracter (SNA) の特徴を示すことが明らかになりました。
2. 生物種の間の相互作用の研究
植食者からの攻撃を受けた植物が特有の匂いを放出して、その植食者を攻撃する捕食者を呼び寄せる現象が広く知られています。面白いことにキャベツでは、モンシロチョウの幼虫に攻撃された場合と、コナガの幼虫に攻撃された場合で反応の仕方が異なります。モンシロの場合はダメージの度合いに応じて匂いシグナルが強くなりますが、コナガの場合には少ないダメージでも強いシグナルを出します。
単一の植物でありながら、攻撃をする植食者およびその捕食者の組み合わせに応じて、シグナルの放出のパターンを変えています。こうしたシグナルの特性の違いを理解するため、相互作用する3種の生物に関して、植物の匂いシグナル放出パターン、植食者の匂いシグナルへの反応の強度、捕食者の匂いシグナルへの反応の強度の3つの戦略に注目し、それらの共進化を理論モデルを構築して調べました。この解析では、植物の状態推移を記述して、それに基づいてそれぞれの生物種の適応度を定式化しました。
植食者が植物を攻撃すると、植物には2つのタイプの変化が生じます。一つは植食者の消費による植物バイオマスの減少です。もう一つは、植食者の攻撃に反応して植物の形質に様々な変化、例えば、成長が顕著になる補償成長や、誘導的な防衛物質の発現などが生じます。植物に起きた変化は、その植物を利用する複数の植食者の間に間接的な影響をもたらします。
消費による資源バイオマスの減少を通じた間接効果は、伝統的に生態学で深く研究され、それらは資源競争やそれに基づく競争排除の理論などに繋がりました。一方、植物の形質変化を介した間接効果の役割は、近年になってようやく注目され始めました。
植物の形質変化を介した間接効果の役割を明らかにするため植物と2植食者の系をモデル化し、植物が非特異的な防衛を誘導する場合に、2つのタイプの間接効果が個体群動態にどのように作用するかを理論的に調べました。それにより、非特異的な誘導防衛の下では植食者の共存が促進され、またその共存状態では1種目の植食者から2種目の植食者に対しては消費による資源減少を介した間接効果が、逆方向には植物の誘導防衛を介した間接効果が働いて、個体数を制御している傾向があることが分かりました。すなわち、植食者の共存状態では、2つの間接効果が同等に重要な働きをしているのです。
3. 協力の進化に関する研究

他個体を助ける利他行動は人間をはじめいろいろな生物で見られます。実は利他行動がコストを伴う場合には「他者を助ける遺伝子」の進化は簡単ではなく、いろいろな条件が必要です。その一つは、助ける個体と助けられる個体の間に血縁関係がある場合に作用する「血縁淘汰」です。
空間構造があって、個体の動きや相互作用が制限される場合には、相互作用個体間の血縁度は高くなるため利他行動が進化しやすくなると思うかもしれません。しかしその場合には血縁者間の競争も強まるために、そのままでは結局は利他行動の進化が制限されてしまいます。
そこで血縁者間の相互作用をより高める要因として、資源をめぐる競争を考えてみました。つまり、資源収奪と利他行動が同時に進化すると仮定するのです。資源収奪と利他行動のそれぞれに影響の到達範囲があるとします。特に資源収奪よりも利他行動の影響範囲が小さい場合、生物の個体の分布に規則的なクラスター構造が自律的に現れ、その中で血縁者間の利他行動が行われることで、血縁淘汰がより効率的に働いて利他行動が進化しやすくなることがわかりました。
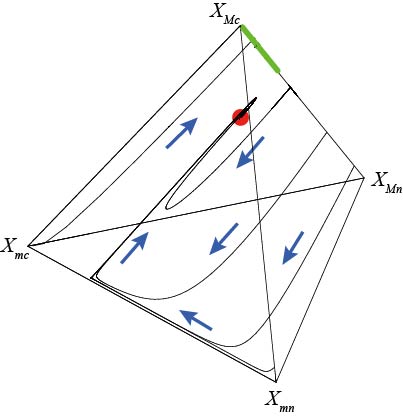
集団内のメンバーが共通の利益を得るために協力関係を構築することもよくみられる現象です。ただ、協力関係の構造によっては、努力を怠って利益だけを享受する「タダ乗り」が集団中に進化して、協力に積極的な個体とタダ乗り個体が共存し、多型状態になってしまう場合があります。こうした進化を適応的ダイナミクスというアプローチで確認すると、集団全体の協力レベルがある値に達したところで「進化的分岐」が起きることによりもたらされることが分かります。
実は協力関係は、2つの取り扱いが組み合わさることで実現されています。各個体が提供する協力への投資の扱いと、投資から得られた成果の扱いです。それらの違いによって例えば、メンバーからの資金を合算して投資をしその成果を分配する場合(資金の合算)や、それぞれの資金を個別に投資しその成果を合算して分配する場合(成果の合算)などの違いが現れます。面白いことに、投資と成果の扱いによって協力の進化の特性、特に協力レベルの多型の現れやすさが異なります。その法則性を理論的に調べました。博士課程の学生だった伊藤公一君との研究です。
4. 細胞内共生体の進化
植物では、集団の中に雄器官(花粉)と雌器官(胚)を同時に持つ個体と雌器官しか持たない個体が共存する場合があり、これを雌性両全性異株といいます。この現象は、しばしばミトコンドリアに生じた雄機能を破壊する突然変異によって引き起こされます。ミトコンドリアは花粉経由では次世代に伝わらないので、花粉の生産をストップさせて胚をたくさん作る方が利益が大きくなるため、そうした進化が起こります。ところが核上の遺伝子にとっては花粉も自身の伝搬に重要な役割を果たすものですから、それを失うことは望ましくありません。そこで雄機能を回復する機能を対抗進化させます。
こうしたミトコンドリアと核遺伝子の間の「ゲノム間闘争」において、自殖(自家受粉)が大きな影響をおよぼします。胚の発生には花粉は欠かせないので、実は雌個体は両性個体からの花粉供給に頼ってしか存続できないのです。そのため、もしも両性個体が自殖ばかりを行って花粉を放出してくれないと、変異体ミトコンドリアの進化は抑え込まれてしまうでしょう。そこで、ミトコンドリアのオス機能抑制遺伝子、核のオス機能回復遺伝子、そして核の自殖率遺伝子の3つの遺伝子の共進化をモデル化し、その過程を解析しました。修士課程の学生だった山岸栄大君との研究です。
